|
||||||||||||
            |

Ivari determináció
| A lényeg: |
A kétnemûség nagyon elterjedt az
élõvilágban, és már nagyon korán
megjelent (az eukarióta egysejtûeknél, mint szinte minden
fontos újítás). Ebbõl azt feltételeznénk,
hogy a mechanizmus, mely által a hím- és nõi funkciók
kialakulnak, szintén nagyon õsi és egységes. Ez
azonban koránt sincs így, az ivari determináció
(ID) útjai nagyon változatosak. Nagy
változatosságot találunk mind az ivardeterminációs
rendszerek mechanizmusaiban, mind az ivari kétalakúság
fenotipikus megjelenésében.
Az ivari funkciók kétalakúságának
kontinuumának egyik végén találhatók azok
a fajok, ahol a mikro- és makrogaméta funkciók két
külön egyedben, szeparáltan találhatók meg
(kétnemû, kétlaki, diöcikus vagy gonochorisztikus
fajok). A másik végletet a hermafrodita fajok mutatják,
itt a két funkció szimultán megvan egy egyeden belül
(mikrogamétát termelõ here és makrogaméta-termelõ
ovárium). A két végállapot között számos
átmeneti alak megtalálható, így pl. androdiöcikus
(hímek és hermafroditák), vagy gynodiöcikus (nõstények
és hermafroditák) fajok.
Annak a megvalósulási módját, hogy egy adott egyed
fenotípusa hím vagy nõstény lesz-e (vagyis az
elsõdleges ivardeterminációs mechanizmusokat) feloszthatjuk
többféle módon. Egy logikus csoportosítás
az alábbi:
1. A megtermékenyítés pillanatában eldõl
az utód neme az ivardeterminációs (és
általában bizonyos kromoszómákon csoportosuló)
gének mûködése következtében. Ezt nevezik
genetikai ivarmeghatározottság-nak (Genetic Sex Determination,
GSD).
2. A zigóta fejlõdése alatt ható környezeti
faktorok befolyásolják az ivart. Ezt nevezik környezeti
ivar-determinációnak (Environmental Sex Determination, ESD).
Itt a megtermékenyülés pillanatában a a genetikai
mechanizmusok még nyitottak, a zigóta fejlõdése
alatt ható környezeti faktorok befolyásolják, hogy
hím vagy nõi nemû lesz az utód. Persze ez az út
is "genetikai", hiszen gének hatnak itt is, így az elnevezések
kicsit megtévesztőek.
3. Populációs tényezõk (ivararány,
pop.denzitás) befolyásolják az egyedek ivarát.
Akár felnõtt korban is ivart válthatnak az egyedek, attól
függõen, melyik ivar bizonyul "kifizetõdõbbnek"
(elõfordul pl. halaknál, fonálférgeknél,
lásd az alternatív szaporodási fenotípusok fejezetet).
A zigóta ivarának kialakításában sokféle ágens vesz részt, ezek a mechanizmusok több szinten fejtik ki a hatásukat, és együttes hatásuk határozza meg az egyed ivarát.
Az elsõdleges nukleáris ID mechanizmusok lehetnek ivarkromoszóma-specifikus transzkripciós faktorok (YX/ZW rendszerek, az ivari kromoszómától függ az utód neme), vagy dózis-dependens transzkripciós faktorok (A:X aránytól függ az utód neme), esetleg hõmérséklet-érzékeny mechanizmusok (a környezei hatástól függ az utód neme).
Ezen elsõdleges faktorok hatását módosíthatják, vagy írhatják teljesen át az ID mechanizmusok másik nagy csoportja, az ivararány-torzítók. Ezek önzõ genetikai elemként viselkedve a saját túlreprezentálásuk érdekében módosítják az elsõdleges ID tarnszkripciós faktorok hatását, és lehetnek nukleáris ivararány-torzítók (B kromoszómák, ivarkromoszóma-kapcsolt gének, vagy ún. komplementer ID faktorok) vagy citoplazmatikus ivararány-torzítók (citoplazmatikus organellumok és citoplazmatikus paraziták, pl.Wolbachia).
1. Ivarkromoszóma-specifikus transzkripciós faktorok
Ezeknél a rendszereknél az ivar-determináns szerepû gének speciális kromoszómákon csoportosulnak, és ezek megléte vagy hiánya befolyásolja elsõsorban az ivart.
XY rendszerek
Azokat az élõlények tartoznak ide, ahol a hímeknek
van két különbözõ, a nõstényeknek
két egyforma ivarkromoszómájuk. Ilyenek pl. az emlõsök.
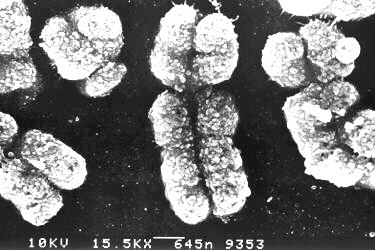
Emlõsöknél a hím a heterokromatikus nem,
XY ivari kromoszómákkal (elektronmikroszkópos
kép) a nõstények XX genotípusúak.
(Heterokromatikus = különböző ivari kromoszómás
ivar, homokromatikus a másik ivar). Aneuploid mutánsok (XXY=hím,
Xo=nõ, nem úgy, mint a Drosophiláknál)
vizsgálatából kiderült: Y kromoszóma a
lényeg az emlős ivar meghatározásában,
nem a kromoszómák aránya. Ezen a kromoszómán
találhatók az ivardetermináns gének (illetve
azok a gének, amely az alapvetõ homokromatikus, nõi
fejlõdésbõl a hím jellegek kialakulásának
irányába "lökik" az embriót).
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Egyéb géneket is találtak a NR-en, melyek fõleg a hímek számára lehetnek elõnyösek. Ez 20 gén (család), szerepük a növekedésben, fogfejlõdésben és spermiogenezisben van (nagy fontosságúak a hím kompetíciós képesség kialakításában, a testméret, harci képességek, spermakompetíció révén).
Növekedés és nemi differenciálódás
Emlõsöknél általános, hogy a hím
embrió gyorsabban és nagyobbra fejlõdik, és nála
hamarabb kapcsol be a testméret-növekedés és az
ivari determinációs mechanizmus, mint nõstény
utódnál. Valószínûleg ez azzal függ
össze, hogy az emlõsõknél hím embrió
az anya testében lényegében nõi közegben,
ösztrogénben úszik, és jó, ha gyorsan és
nagy ivari jellegek alakulnak ki, hogy elnyomják a feminizáló
hatást (gyors növés=stabil here). Ez a nõsténynek
nem gond, mint ahogy madaraknál sem, hiszen õk nem az anyában
fejlõdnek.
ZW rendszerek (nõstény a heterokromoatikus, hím a homokromatikus ivar)
Erre jó példa a madarak. Itt a nõstény a hetetrokromatikus ivar, (ZW ivari kromoszómákkal), a hímek ZZ genotípusúak. Nem tudni, hogy W kromoszóma a lényeges, vagy a Z kromoszómák száma. Aneuploid mutánsok nem ismertek, azok alapján meg lehetne mondani. Itt is van pszeudoautoszomális régió a Z kromoszómán és szintén kevés gén található rajta.
Madár és emlõs (és
egyéb) ivari kromoszómák nem homológok, más-más
autoszómákból, függetlenül fejlõdtek
ki. Pl. az emlõs X és madár Z kromoszómákon
csak egy közös gén van (OTC), de ez akár véletlen
is lehet, viszont a Z az emlõs 9. autoszóma nagy részével
homológ.
|
Az ivari kromoszómák kialakulása és evolúciója
A nemrekombináns részek lényegében
klonálisan terjednek. Olyan gén, amely elõnyös
egyik, hátrányos másik ivarnak, jobban elterjedhet |
Az ivardetermináns géneket hordozó speciális kromoszómák száma dönt az ivar kialakulásáról (autoszóma:.ivarkromoszóma aránya számít).
A Drosophila legyeknél
is a X kromoszóma-autoszóma arány számít
(Y kromoszóma van, de nincs ivardetermináns szerepe).
XX: nõstény, XY: hím (aneuploid XXY mutáns
is nõstény, míg az Xo mutáns steril hím,
nem úgy mint emlõsökben, ebbõl látszik,
hogy Y nem elsõdleges az ivardeterminációban). Az
X -ek tartalmazzák a nõt determináló géneket,
az autoszómák a hímet. Ha a genom két X-et
tartalmaz, bekapcsol négy X-en lévõ numerátor
és egy autoszomális denominátor gén. Ezek
szabályozzák a Sex-lethal (Sxl) gén mûködését,
ami a nõsténnnyé fejlõdéséhez
kell (sepcifikus tra mRNS splicing és dsx által
megvalósítva a nõstény fenotípust).
Coenorhabditis elegans hengeresférgeknél
is X kromoszóma és autoszómák aránya
számít a nem meghatározásában, de itt
nincs Y kromoszóma. XX genotípus hermafrodita, Xo genotípus
hím lesz. Komplex génsorok együtthatása határozza
meg a nemet, az X-ek számától függõen
(xol-1 gén esszenciális a folyamatban). Ez a faj
amúgy androdiöcikus (hímek és hermafroditák
vannak), és a hermafroditák képesek önmegtermékenyítésre,
míg a hímek csak spermiumot termelnek.
Ez esetben valamilyen környezeti hatástól függõen (ez általában a hõmérséklet) lesz hím vagy nõstény a magzat, vagyis a megtermékenyüléskor még nyitott annak lehetősége, hogy milyen utód fejlődik ki.
Ilyen rendszert találtak már az említettek kívül kerekesférgeknél, hengeresférgeknél, gyûrûsférgekben, és növényekben is. A környezet hatása taxononként változó, van ahol meleg hatására inkább hímek, hidegebb környezetben nõstények lesznek, de fordítva is lehetséges. A mocsári teknõsnél pl. 25 fokon minden tojásból hímek, 30 fokon pedig nõstény utódok kelnek ki. A köztes hõmérsékleteken vegyesek az almok. A pontos mechanizmus és a ható gének még nem ismertek.
| A Chelidra serpentina teknõsnél alacsony
és magas fészekhõmérsékletnél
nõstények, míg közepesnél fõleg
hím utódok születnek. |
{kind=link}
Hormonkezelés (tesztoszteron, ösztrogén) elfedi TSD hatását, ezért (legalábbis hüllõknél) ösztrogén/tesztoszteron-szint növekedés (bizonyos hõmérséklet által kiváltva) lehet a molekuláris mechanizmusa az ivarcsinálásnak.
A jelenség adaptív voltára számos hipotézis szolgál magyarázattal. A leginkább elfogadottak szerint a fejlõdési hõmérséklet befolyásolja a kikeléskori fenotípust, és ez különbözõ mértékben befolyásolja a két ivar fitnessét. Ha pl. a fejlõdési hõmérséklet kedvez a növekedésnek (nagyobb utódok születnek, mint kisebb hõn), akkor ezen a hõmérsékleten érdemesebb olyan nemû utódnak kialakulnia, mely jobban tud profitálni a megnövekedett méretbõl vagy gyorsabb növekedésbõl.
|
Alligátoroknál melegebb hõmérsékleten több hím kel ki, és a nagy testméretû hímek skeresebbek a háremtartás növekvõ sikeressége miatt. A kalászhalak nõstényei annál nagyobb arányban fialnak hím utódot, minél melegebb a víz (akár a laborban is). Valószínûleg a hideg vízbõl arra következtethetnek, hogy a szezon még korai szakaszban van, a kikelõ utódnak még sok ideje van fejlõdni. Mivel a nagyobb testméret a nõsténynek kedvez (nagy test, sok ikra, nagy szaporodási siker), hidegben több nõstény kel ki. |
{kind=link}
4. Haplodiploidia
A hártyásszárnyúak nemi determinációja
alapvetõen az arrhenotokia, a megtermékenyített petékbõl
diploid nõstények (és dolgozók), a haploidokból
hímek kelnek ki. A pajzstetveknél pont fordítva van,
ott a diploid peték lesznek hímmé (teliotokia).
Ebben az ID rendszerben az ivararány primer befolyásolása (utód nemének eldöntése) könnyen lehetséges, a tojást rakó állat dönthet az utód nemérõl (megtermékenyíti-e a petét lerakás közben vagy nem). Más ID rendszerû állatoknál még nem találtak bizonyítékot primer szabályozásra.
Növények
A növények ebbol a szempontból nagyon változatosak.
Pl. a virágos növények 72 %-a hermafrodita (egy virágon
belül hím és noivarsejtek) 7 %- a monoöcikus (vagy
csak hím, vagy csak noi virágok, de egy egyeden vegyesen),
17% átmeneti (hermafrodita és monocöikus keveréke)
és csak 4 % dioöcikus (kétlaki, külön hím
és no virágok, és egy egyeden csak egyik félék,
ugyanúgy, mint a kétnemu állatoknál). A kétlakiakon
belül is változatos a determinációs rendszer
,akár csoportokon belül is nagy varianciát mutathat.Van
példa XY rendszerre is, mint az emlosöknél. Az Y ivari
kromoszóma itt aY bibefejlodést gátolja és
a porzó fejlodését indítja be.
|
Egy részletesen vizsgált növényi XY rendszer a Silene szegfûnemzetségé. Itt is morfológiailag különbözõ X és Y kromoszómák vannak, mint pl. az emlõsöknél, van rekombinálódó kicsi pszeudoautoszómális régió (PAR), klonális NR. A rekombináció szupressziója itt is hasonlóan zajlott, mint emlõsökben, fokozatosan növelve a nem-rekombinálódó részek arányát a kromoszóma-páron. (Az X és Y-n is megtalálható közös, de már nem rekombinálódó gének divergenciája nõ a PAR-tól távolodva.) A két kromoszóma kb. 10 millió éve kezdhetett morfológiailag elkülönülni, vagyis jóval fiatalabb, mint az emlõsök ilyen rendszere. |
Az ivardeterminációs utak evolúciós dinamikája
Az ivardetermináns mechanizmusok behatóbb, filogenetikai rendszerben történõ vizsgálata az ID-rendszerek és gének meglepõen gyors evolúcióját mutatják. Adott gének nem feltétlenül vesznek részt a rokon fajoknál az ID-ban, és közeli rokonok alapvetõ ID mechanizmusokban különbözhetnek egymástól. Akár fajon belül is lehet variancia (pl. Xiphophorus maculatus halnál XY és ZW rendszerû populációk is ismertek)
Az emlõs ivardetermináns faktor, az SRY
(és az egész Y kromoszóma) is gyors evolúciót
mutat:
Az SRY tipikus Y gén, autoszómáról jött, gyors az evolóciója, gyorsan változik, gyorsan el is tűnhet. A kromoszóma kezdeti ivarkromoszómává válását elindító TDF nem az SRY volt, hanem valami más faktor. Az SRY őse a SOX3 gén lehetett, ami főleg idegrendszer-fejlődésben szerepel manapság, ez módosulhatott ivardetermináns génné az Y-ra kerülve, most már ez a TDF. Az Y kromoszóma minden emlősben megvan, madárban és hüllőben nincs, kb. 310 - 170 millió évvel ezelőtt jelent meg. SRY viszont Monotrematákban ban nincs még meg, de erszényes és méhlepényes emlősökben van, ergo az SRY kb. 170-130 millió éves, ekkor módosulhatott SOX3-ból.SRY tipikus Y génként gyorsan változik, a Mus genusban SRY mósosult Sry-vé, az Ellobius genus (vakondpockok) néhány fajában Sry már eltűnt. E. fuscocapillusban van Y kromoszóma és Sry, de az E. lutescensnél: (hím és nõ is Xo, nincs Sry) és az E. tancreinél (a hím és nõ is XX, nincs Sry) eltűnt. |
A Silene szegfûknél a nemzetségen belül is figyelhetõ meg változatosság, az XY renszdszerû kétivarú fajok mellett (pl. Silene latifolia, Silene dioica, lásd korábban) hermafrodita fajok is vannak, (pl. Silene vulgaris, Silene diclinis).
A Caenorhabditis férgeknél az androdiöcikus rendszer egymástól függetlenül alakult ki a C. elegans és a közel rokon C. briggsae esetében nem önmegtermékenyítõ, diöcikus õsökbõl. Ráadásul az ID-ben fontos xol-1 gén igen gyors szekvencia-divergenciát mutat, a C. elegans és C.briggsae fajokban csak 22%ban identikus, míg környzõ, nem ivardetermináns gének sokkal kevésbé gyors változást mutatnak.
A Drosophila legyek és egy közel
rokon gyümölcslégy genus esetében is azt találták,
hogy az ID utak jelentõsen különböznek. Míg
a Drosophila-ban az: X:A arány az Sxl génen keresztül
vesz részt a tra mRNS splicing-ban a nostény fenotípus
kialakításához (lásd elõbb), a Ceratitis
nemzetségben az: Slx nem ivardetermináns gén,
a tra-reguláció autonóm.
Néhány további példa az elsõdleges
nukleáris ID rendszerek változatosságára:
Hüllõkben sokféle mechanizmus
van, ezek nem is köthetõk egyes taxonokhoz. Krokodiloknál ESD, teknõsöknél
ESD és GSD (XY és ZW rendszer is), kígyóknál
GSD (ZW rendszer), gyíkoknál pedig ESD és GSD (XY és
ZW is).
Kétéltûeknél is
vegyes, van ESD és GSD is (XY és ZW).
Halaknál is sokféle ESD és
GSD (XY és ZW).
Lepkéknél genetikai (ZW rendszerû)
a determináció.
Ivararány-torzító SD mechanizmusok
Eddig az elsõdleges ID mechanizmusokat néztük, át, és láttuk, mennyire sokféle módon valósulhat meg a kétnemûség (aminek pedig mindenhol ugyanaz a lényege), és ráadásul ez a sokféleség is dinamikus változásban van. Az ID mechanizmusok másik nagy csoportja, az ivararány-torzítók ezen elsõdleges ID faktorok hatását módosíthatják, vagy írhatják teljesen át.
1. Nukleáris ivararány-torzítók
Komplementer ivar-determináció (Complementary Sex Determination,
CSD)
Ez a mechanizmus eddig csak hártyásszárnyúaknál
ismert, és ezek elsõdlegedes ID-jét (haplodiploidia)
nagymértékben módosítja. Ez esetben egyetlen
egylókuszos gén alléljaitól függ az utód
neme, nem a haplodiploidiától. A diploid heterozigóták
(AjAi) nõstényekké, a diploid homozigóták
(AjAj vagy AiAi) és haploid hemizigóták (Aj vagy Ai)
pedig hímekké fejlõdnek.
A legkülönbözõbb Hymenoptera fajokban ismert
a jelenség, így euszociális hangyáknál,
méheknél és magányos parazitoidoknál
is. Az ilyen diploid hímek diploid spermát termelnek, és
effektíve sterilek. A steril hímek a populációnak
nagy genetikai terhet jelentenek, melynek mértéke nõ
a beltenyésztéssel, hiszen ekkor a homozigóta egyedek
aránya is növekszik. A CSD lókuszon az allélek
száma fajonként különbözõ lehet, átlagosan
9-20, ennek megfelelõen 11 és 5 % között mozog a
diploid hímek populáción belüli aránya.
A CSD nagymértékben befolyásolja mind a szociális
viselkedést, mind a tényleges és az effektív
ivararányt, valamint a párzási rendszerek kialakulását.
|
Ha a nõstény hetrozigóta (AjAi) és más allélú haploid hímmel (Ak) párzik, a születõ diploid utódok mind nõstények lesznek, ez esetben nem jelenik meg a diploid hímek terhe. Ha a heterozigóta (AjAi) nõstény azonos allélú haploid hímmel (Aj vagy Ai) párzik, a diploid utódok fele steril hím lesz, ha pedig diploid hímmel, akkor a megtermékenyített peték mindegyike életképtelen, a nõstény ez estben csak megtermékenyítés nélküli haploid hímeket tud produkálni. Az ilyen párosodott nõstények funkcionálisan termékenyítetlennek tekinthetõk. Nem szociális hártyásszárnyúaknál a CSD nagy hatással lehet a populáció növekedésre és az ivararányra. A sok steril hím miatt az effektív ivararány erõsen a nõstények felé mutat, a szaporodóképes egyedek nagy része nõivarú. Mivel a diploid hímekkel párosodó nõstények csak haploid hím utódot képesek létrehozni, szelekciós nyomás van a nõstények felé, hogy nõi ivar felé tolják el a születendõ utódok nemét, ezzel kompenzálva a sok hímet. A társas fajoknál a dolgozók miatt összetettebbek a CSD hatásai. A dolgozók genetikailag nõstények. A diploid hímek produkciója csökkenti a dolgozók számát is, nagy lassulást okozva a pop.növekedésben. |
| Sok
fajnál alakult ki valamiféle szociális viselkedésmód
a diploid hímek okozta hatások kompenzációjára.
Apis mellifera dolgozók diploid lárvákat
felismerik és megeszik, Melipona méhek dolgozói diploid
hímmel párzott királynõjüket (a kolóniát
egy nõstény alapítja, amely egyetlen hímmel
párzik) megölik, és a testvérei közül
emelnek egy új királynõt, Formica fajoknál a dolgozók eliminálják
a diploid hímeket. Hatékony védekezés
lehet még a beltenyésztés elkerülése
diszperzióval. A Solenopsis invicta tûzhangyánál az eredeti élõhelyén, Argentínában 86, a behurcolt USA-i populációkban 10-13 CSD allél meglétét mutatták ki. Az allélszám-csökkenés valószínûleg bottleneck hatás miatt van. A sok CSD allél miatt Argentínában nagyon kevés diploid hímet találtak, az USA-ban nagyon sokat (nagy a homozigócia esélye). |
{kind=link}
{kind=link}
{kind=link}
2. Citoplazmatikus ivararány-torzítók
Bizonyos sejtorganellumok (mitochondrium, kloroplaszt) és egyéb genetikai elemek, citoplazmatikus paraziták uniparentálisan örökítõdnek (általában maternálisan, lásd elõbb). A nukleáris gének érdeke nagy populációban és pánmixisben az 50% hím-50% nõ arány, de a citoplazmatikus elemek génjeinek érdeke a 100% saját ivar (másik ivar zsákutca, nem képes õket örökíteni) létrehozása. Egy ilyen rendszerban szelekciós nyomás van arra, hogy bizonyos faktorok önző genetikai elemekként viselkedjenek, és befolyásolják az ivararányt az õket örökírõ nem javára. Ez a konfliktus igen nagymértékben befolyásolhatja az ivararányt (citoplazmatikus ivararány-torzítók). Hatásuk lehet feminizáció (zigóta fejlõdését nõi irányba tolják), hímsterilitás vagy hímölés, (hím zigótát elpusztítják, vagy sterilizálják) partenogenetizálás és meiotikus sodródás okozása. A lényeg mindegyikben az, hogy a nõi gamétát (amiben õk örökítõdnek) termelõ ivar felé tolják el az ivararányt.
Az ivararány-torzító genetikai elemek szelekciós nyomást gyakorolnak olyan nukleáris genetikai elemek evolúciójára, melyek ezeket a torzítókat szupresszálják, hasonlóan az uniparentális öröklõdésben (lásd előbb) részt vevõ genetikai elemekhez (fegyverkezési verseny). A két szelekciós erõ kölcsönhatása nagymértékben befolyásolhatja, vagy teljesen meg is változtathatja az eredeti ID mechanizmusokat.
![]()
Armadillidium vulgare szárazföldi ászkáknál ismertek olyan Wolbachia baktériumok, amik a faj ivardeterminációs rendszerét alapvetõen megváltoztatják. Ezek a baktériumok intracelluláris,
anyai ágon örökítodõ szimbionták/paraziták.
Számos rovar, rák, fonalféreg lehet a gazdája.
Ismert hatásuk a gazdára sokféle lehet: A Wolbachia-mentes
populációkban a hímek ZZ, nõstények
ZW ivari kromoszómájúak (nõi heterokromácia).
A fertõzött populációkban a baktérium
a ZZ genotípusú hímekbõl nõstényeket
csinál, melyek szaporodóképesek, és
örökítik a baktériumot. Sok populációban
szinte kizárólag ZZ nõstények vannak,
melyek tartalmazzák az Wolbachiát, így a W kromoszóma (az
eredeti nõstény-determináns faktor) kihalásra
van ítélve (nõ és hím is
ZZ). Mivel szinte minden utód nõstény lesz
(ZZ genotípus, de baktérium nõsténnyé
teszi), a hímek nagyon ritkák, és nagy
a fitness-elõnyük (frekvenciafügõ szelekció),
ezért nagy szelekciós nyomás van a baktérium-mentes
ZZ hímet csinálni tudó (mentes) nõstényekre
(ZW). Meg is jelentek az Wolbachiát gátló autoszómális
szupresszor gének, amik mentes egyedeket eredményeznek. |
{kind=link}
Humán ivarfejlõdés
1. Kromoszómális nem
A zigóta nemi
kromoszómáit jelöli.
Embernél genetikai determináció van, a
zigóta genomja (XX vagy XY) predesztinálja a
születõ utód nemét (van kivétel,
lásd késõbb).
2. Gonadális nem (a
magzati fejlõdés 6.
hététõl)
Az elsõdleges ivarszervek (here
és ovárium)
kialakulása után beszélünk hím
és nõi gonadális nemekrõl.
Génhatásra zajlik a folyamat, végeredménye
a here vagy a petefészek kifejlõdése. Mindkettő
ugyanabból a gonádtelepből fejlődik, mely az
õsvese alatt helyezkedik
el, és az ötödik hétig indifferens, hím
és női egyedben megkülönböztethetetlen. Ekkor az
õsivarsejtek bevándorolnak a telepre, és a
bekapcsoló gének hatására a telep
differenciálódni kezd.
A ható gének egyszerûsítve:
Y kromoszóma SRY régiója,
a TDF (Testis determinating factor)
X kromoszómán: TIF (Testis
inhibitor) gén (a here fejlődésének indulását
akadályozza meg)
O gén (ováriumot csinál kortikalizációval
a telepbõl)
T gén (herét csinál medullarizációval a
telepbõl)
Az embrió fejlõdésének hatodik hetén indul be SRY, TIF gátlódik, a T gén indulni tud, medullarizáció, és here lesz. Ha nincs Yp régió (női az egyed és nincs Y kromoszómája, vagy hibás az Yp gén), akkor az O gén indul, kortikalizáció, ovárium lesz, here nem alakul ki. Látható, hogy az eredeti ivari irány a női (homokromatikus), és a meglévő Y kromoszóma hatására lesz hím a delikvensből.
3. Genitális nem (12.
héttõl)
A differenciálódó ivari traktusok alapján beszélünk
hím vagy nõi genitális nemrõl. A kialakult here
által (hím a gonadális nem) termelt csecsemõkori
tesztoszteron hatására alkulnak ki a nemi különbségek.
A genitáliák fejlõdése (is) alapvetõen
mindig a homokromatikus (női) nem irányába halad. A csecsemõkori
tesztoszteron "löki" hím írányba a genitáliák
fejlõdését. (Müller-csõ elsorvad, Wolf-csõ
differenciálódik mellékherévé, ondóvezetővé,
pénisszé és herezacskóvá). Ha nincs here,
akkor nincs tesztoszteron, a Müller-csõ fejlõdik tovább,
nõi irányba halad a zigóta (petevezetõ, méh,
vagina és vulva alakul ki a Müller-csõbõl).
4. Pszichoszomatikus
(társadalmi) nem
Ez az, aminek az egyén tartja
magát
(neveltetés, kulturális
hatások, stb. erõsen befolyásolhatják)
Turner-szindróma (X0)
Klinefelter-szindróma (XXY)
"Szupernõ" (XXX)
Euploidia esetén (ivari kromoszómák kiosztásával nincs baj):
1. Sex reversal
Az egyed kromoszómális neme stimmel (XX - nő), de az ivar-determinációban ható génszakaszok nem a megfelelő ivari kromoszómán vannak: (rekombináció van, Yp szakasz X-re kerül). Ekkor a kromoszómálisan női egyednél ugyanúgy beindulnak a "hímcsináló" folyamatok (here, hím genitáliák lesznek), végeredményben teljes értékû férfi lesz az eredetileg női zigótából.
Ha az XY-os férfinak hiányzik
az Yp szakasza az Y-ról, akkor is sex reversal van, teljes értékû
nõ lesz a fenotípus (TIF gén hat, nincs here, nincs
tesztoszteron, minden női irányba
halad)
2. Ha tesztoszteron-hatás éri
a fejlõdõ női magzatot (gyógyszer, mellékvesei
enzimdefektus, stb.) Ovárium kialakul, de az androgén hatások
miatt hím irányba megy el a fejlõdés. A hormonhatás
idõpontjának függvényében hím genitáliák
lesznek (herezacskó vulva helyett, pénisz csikló helyett,
stb.). Minél korábban éri a tesztoszteron-hatás,
annál jobban torzul az eredeti nõi fejlõdési út.
3. Álhermafroditizmus: (tesztikuláris
feminizáció) Csecsemõkori tesztoszteron hiánya
miatt alakul ki férfiaknál. Ekkor a csecsemõkori here
kialakul, de valami gubanc miatt nem termel tesztoszteront, a genitáliák
nõi irányba fejlõdnek (Müller-csõ), nõként
él nemi életet, de petefészek hiányában
steril.
