|
||||||||||||
            |

A szexuális szaporodásnak van néhány alapvetõ és általános jellemzõje, melyek az ilyen módon szaporodó élõlények túlnyomó többségében megtalálhatóak. Ezek:
1. Meiotikus ivarsejtképzõdés
2. Haploid-diploid ciklusok váltakozása
3. Rekombináció és crossing over
4. Genomfúzió(szingámia)
5. Bináris szexuális rendszer (két nem)
6. Uniparentalitás (a citoplazmatikus organellumok csak az egyik
szülõ általi örökítése)
7. Anizogámia (a "hím" és "nõi" ivarsejtek
eltérõ mérete)
Temészetesen ezek a tulajdonságok folyamatosan jöttek
létre a törzsfejlõdés során, és
vezettek a mai kép kialakulásához. A következõkben
ezen fontos újításokat vesszük sorba, megvilágítva
azokat az elõnyöket, melyeket alkalmazásuk jelentett.
Az eukarióta szex annyiban eltér a prokarióta mechanizmusoktól,
hogy mások a sejtes mechanizmusok, és a genetikai állomány
sokkal nagyobb részére terjed ki a rekombináció.
Molekuláris szinten azonban megfigyelhetõ bizonyos folytonosság
a rekombinációs és DNS-javító enzimekben.
Ezek ellenére a mai modern szexualitáshoz vezetõ
lényeges újítások mind az eukarióta
egysejtû élõlényekben játszódhattak
le, kb. 1 milliárd éve (prokariótákban is
van valamilyen szintű génkicserélődés,
lásd aszimmetrikus szexuális folyamtok). Valamiképpen
mindegyik előnyt jelentett az adott ősi populációban
az újtásokat nem "alkalmazó" egyedekkel szemben,
és ezért elterjedhettek, és máig megőrződtek.
A haploid-diploid ciklusok elõnyei
Ilyen ciklus már a rekombináció feltalálása elõtt is létezhetett. Kezdetben történhetett endomitózissal (Barbulanympha ostorosok, Saccharomyces élesztõ ma is) a haploid-diploid ciklus váltakozása, összeolvadás és crossing over nélkül nélkül. (Ezt bizonyítja, hogy a kromoszóma-párosodás és a crossing over szabályozása különálló géncsoportok által történik).
Ha a ploidiaszint meghatározza az
ellenállóképességet és a környezet
változékony, akkor ökológiailag elõnyös
lehet a ploidia-váltakozás, melyben az egyes formák
a különbözõ környezethez való alkalmazkodást
jelentik (pl. a DNS-károsodás mértéke váltakozik
a korai légkör oxidatív gyökeinek hatása
erõsségének függvényében). Olyan
élõlényeknél, ahol nincs mitokondrium és
peroxiszóma (pl. Archezoáknál) nagy a szabadgyökök
kárósító hatásának veszélye,
nagyon elõnyös lehet egy ellenálló genomi állapot.
Számos tény mutatja, hogy a diploid formák ellenállóbbak
lehetnek (pl. élesztõgombák).
Ha a diploidia ilyen nagy elõnyökkel jár, akkor miért
kell a haploidiára visszatérni?
1. Haploid állapotban fele akkora a genetikai teher (diploidokban dupla a genomra vonatkoztatott mutációs ráta)
A genomfúzió (szingámia) elõnyei
Láttuk, hogy a haploid-diploid ciklus bizonyos körülmények között elõnyös lehet, de ha ezt lehet edomitózissal is biztosítani, melyhez nem kell egy másik egyed sejtjével fuzionálni, akkor miért (szinte) kizárólagos mégis a genomok fúziójával való diploidizálódás?
Hickey és Rose alternatív hipotézise (1988) szerint a fúziót lehet, hogy endoszimbionta parazita vagy egyéb önzõ genetikai elem váltotta ki, saját horizontális terjedését segítendõ (pl.Physarum nyálkagomba mitokondriális mif+ plazmidja vagy E. coli F-plazmidja, lásd a szexualitás előnyeinek taglalásánál).
{kind=link}
{kind=link}
A crossing over haszna
A crossing over és a rekombináció valószínûleg a szingámia és az egylépéses meiózis kifejlõdése után jelent meg, hiszen endomitózisnál fölösleges lenne (homozigóták a haploid állapotok).
Populációs szinten jelentkezõ elõnye a crossing overnek, hogy a gyorsabb alkalmazkodást és a káros mutációk okozta fitness-csökkenés mérséklését elõsegíti (bõvebben lásd az elõzõ fejezetben). Egyedi szintû elõnyök nélkül azonban nem alakulhatott volna ki a crossing over, így ilyeneket is fel kell tételeznünk. Ezek lehetnek:
1. A szinergeta káros mutációk szelektív nyomással vannak a rekombináció kialakulására, mely lehetõvé tesz a szétszeparálásukat (külön nem olyan károsak).
2. Ha irányító
szelekció hat az adott többlókuszos
tulajdonságra, akkor a szelekció a
rekombinációt segítõ génnek is
kedvez (pl. parazita nyomásra adott jelleg variábilisabb
lesz, ezáltal elõsegítve a menekülést
a parazita elõl).
Ez a két szelekciós hatás (vagy az egyik)
vezethetett a crossing over kifejlõdéséhez. Ez a
lépés nagyon fontos következményekkel
járt, de kialakulásához minden
szükséges tényezõ (meiózis,
enzimek) adva volt már korábban.
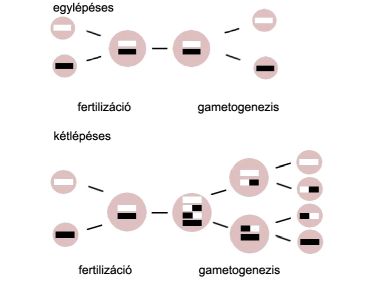
A kétlépéses meiózis elõnyei
Számos protista fajnál találtak egylépéses meiózist (Archezoa, Dinozoa, Sporozoa, Parabasalia). Ekkor a fúzió után a homológ kromoszómák premeiotikus duplikáció nélkül szétválnak (és nem tudni, van-e rekombináció).
Csak a kétlépéses meiózisnál
van premeiotikus megkettõzõdés (metakarióta
újítás, a genom előbb megduplázódik,
hogy aztán két lépésben megfeleződjön),
ami lehetõvé teszi:
1. A testvérgyilkos allélek leszerelését (olyan
önző genetikai elem, ami saját terjedését
elõsegítendõ elpusztítja azt a testvérgatétáját,
amiben nincs jelen), ez esetben a kétszeri osztódás
miatt 50%-ban a saját allélt tartalmazó gamétát
pusztítja el, így nem jelent reproduktív elõnyt.
2. A kétszálú DNS premeiotikus duplikációnál
a legutóbbi mitózis óta keletkezett DNS-hibák
javítását (sima mitóziskor is történik
javítás). A duplikációval az esetleges javíthatatlan
egyszálú hibát tartalmazó DNS javítható
kétszálúvá alakul.
A gaméták differenciálódása
Ma is fellelhetők olyan élőlények, melyek izogámok (ivarsejtjei, ill. maguk az egysejtű lények nem különböznek, pl. Clamydomonas algák). A túlnyomó többség azonban differenciált gamétákkal bír. Miért?
Hoekstra ('87)modellje:
1...........................................................................................2.
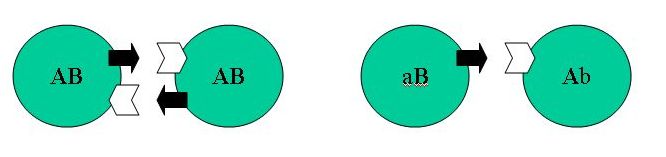
2. Kieséses mutációkkal
Ab és aB típusok is megjelennek (Ab csak aB-vel tud fuzionálni).
AB típus elõnyben van:
kétszeres esélye van a szaporodásra (Ab-vel
és aB-vel is képes). Szelekciós erõ
hat A és B kapcsoltságra, mivel ab genotípus senkivel sem képes
szaporodni.
Ha az eltérõ típussal való párosodás
valami egyéb elõnnyel jár, akkor mégis elterjedhetnek
a Ab és aB típusok, a kezdeti AB típus kiszorítható, a differenciálódás
fixálódhat. Ilyen elõny lehet a heterozigócia
(hibrid vigor), a genetikai konfliktusok csökkentése, zigóta-túlélés
növekedése vagy a pártalálás nagyobb
esélye. Mindegyikre számos hipotézis született,
ezek közül tekintünk át néhányat.
Uniparentális öröklõdés
Genetikai (vagy intragenomiális) konfliktus alatt azt értjük, ha ugyanazon egyedben különbözõ genetikai elemek érdekei ellentétesek, terjedésük más gének terjedésének rovására történik.
![]()
Az eltérõ szülõktõl származó
citoplazmatikus organellumok (mitokondrium, kloroplasztisz, stb.) a zigótában
konfliktusba kerülhetnek egymással. Mivel ezek külön
génállománnyal rendelkeznek, külön szelekció
alatt állhatnak, eltérõek lehetnek az "érdekeik".
Egy mutáns mitokondrium allél pl., amely a másik
szülõbõl érkezõ mitokondriumokat elpusztítja,
elõnyben van a továbbörökítésben,
és elterjedhet. Ha két ilyen mt egy utódba kerül,
az káros hatással járhat az utódra (pl. mindkét
mt eliminálódik, akkor nincs mivel lélegezni, a zigóta
elpusztul). Emiatt konfliktus alakul ki az önzõ mitokondrium
(érdeke a mindenáron való továbbadódás)
és nukleáris gének között is (érdekük
a konfliktus leszerelése). Emiatt szelekciós nyomás
van a konfliktus leszerelésére (nukleáris gének
irányításával). Az efféle konflikusokat
semlegesíteni képes élõlények mérsékelhetik
az önzõ genetikai elemek káros hatásait, és
ezáltal szelekciós elõnyt élvezhetnek. vázlatos
rajz a folyamatról
A citoplazmatikus konfliktus leszerelésének két alapvetõ
módja van:
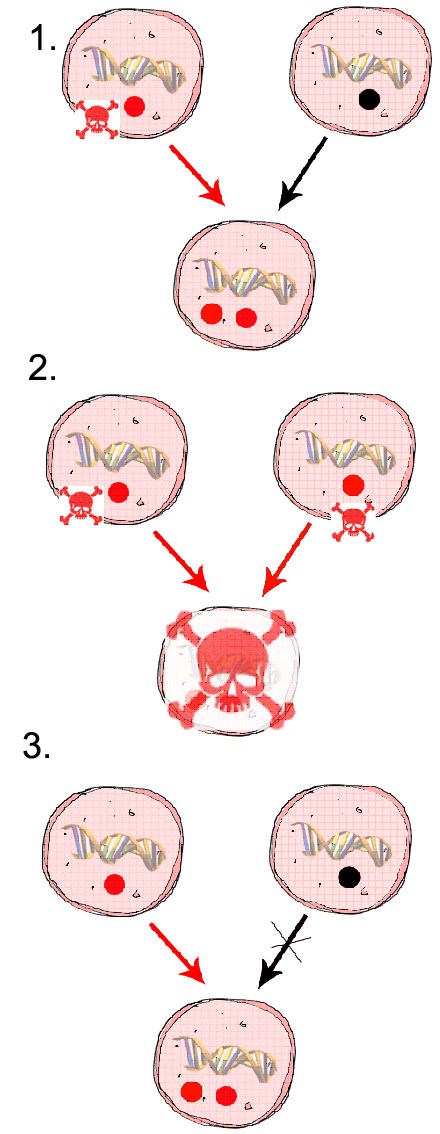{kind=link}
1. Az organellumok fúzió
utáni gyors szegregációja a zigótában
2. A citoplazmatikus organellumok uniparentális
öröklõdése
Ez az uniparentalitás nagyon változatos formában valósulhat meg az egyes fajokban:
A meiotikus osztódás során az ivarsejtekbe nem (vagy csak egyféle) organellum kerül. (Pl. Cilindrocystis zöldalgák).
Az ivarsejtben az organellumok vagy organellumgének degradálódnak (pl. Temnogyra zöldalgák).
2. Fúziókor
A zigótába csak az egyik ivarsejt organellumai jutnak.
| Nyitvatermõknél a kloroplasztiszt a hím
gaméta, a mitokondriumot a nõi gaméta viszi
a zigótába (ezért nem is lehetnek szûznemzõk)
Páfrányoknál a spermium citoplazmája nem jut az utódba |
3. A zigótában
fúzió után
Bizonyos egysejtûeknél
a heteroplazmikus (mindkét szülõtõl
vannak organellumai) zigótában a
különbözõ eredtû
szervecskék szegregálódhatnak, és
néhány osztódás után homoplazmikus
(csak egyféle szülõtõl vannak organellumai)
sejtek keletkezhetnek. (pl. Saccharomyces élesztõ).
{kind=link}
{kind=link}
| Chlamydomonas algákban izogámia
van, + és - párosodási típusokkal. A
zigóta mindkét féle szükõtõl
örökli a mt-t, de csak + típus marad meg (kölcsönös
nukleázos emésztés van, és + mt jobban
bírja, esetleg a - típusban leszerelõ gének
hatnak). |
{kind=link}
Az uniparentalitást biztosító különféle mechanizmusokat szinte minden esetben nukleáris gének irányítják. Az egyes típusok azonban nem köthetõk taxonokhoz, a magvas növények fajai pl. minden említett mechanizmusra szolgáltatnak példát.
| Emlõsöknél pl. a spermium nagyon kevés mt-t juttat a zigótába, fúzió után pedig még ezek is degradálódnak. |
A bináris nemi rendszer kialakulása
Ahol van szexualitás, ott szinte mindig két párosodási típus (ivar) fordul elõ. Ez a bináris rendszer két módon alakulhatott ki:
1. A párosodási típusok hiányából
alakult ki kétféle típus.
2. Sok párosodási típusból redukálódott
kettõvé.
A elsõ lehetõséget sok elmélet magyarázza, mindegyik a citoplazmatikus konfliktusok szerepét hangsúlyozva a folyamatban. A szex vagy bináris (vagyis két ivar van), vagy nem létezõ, de sohasem sokszoros (azt nem ivarnak, hanem inkompatibilitási rendszernek hívják, lásd késõbb) és ez nagyon korrelál a citoplazmatikus fúzió meglétével és hiányával.
Az ezt magyarázó két fõ elmélet:
1. A bináris rendszer a zigótán belüli citoplazmatikus (CP) organellumgének konfliktusai miatt jött létre (Hurst-Hamilton modellje)
| 1. Kezdetben izogám, Clamydomonas-szerû lények, párosodási típusok nélkül. Minden gamétában vannak saját genommal rendelkezó cp gének, ezek passzívak 2. Egy cp+ gén, amely a másik szülõbõl származó cp-ket elpusztítja. A továbbadódásra így kétszeres esélye van, elterjedhet a populációban, amíg a költségei nem lépik túl a kétszeres elõnyt. Költség lehet az eliminálás miatt, (végsõ esetben, két cp+ gaméta találkozásakor a kölcsönös elimináció miatt a zigóta károsodása, akár halála). 3. Egy nukleáris
szupresszor gén, ami a saját gamétájában
meggátolja a cp+ gént az eliminálásban, és
a cp- gént megvédi az eliminálástól
megjelenik. Két nem-szupresszor
génes gaméta találkozása magában
hordozza valamelyik (mindkét) clp eliminációjának
veszélyét. A szupressziónak viszont van valamilyen
költsége. 4. Egy nukleáris válogatós (choosy) gén, amely felismeri és preferálja a saját hordozógamétájától eltérõ szupresszor-típussal történõ fúziót, elterjedhet, mert optimális párzást biztosít. 5. Ilyen feltételekkel, random párosodást feltételezve a rendszer minden esetben bináris irányba mozdul el, ahol 1:1 a párosodási típusok aránya. |
2. A bináris rendszer a citoplazmatikusan terjedõ paraziták miatt jött létre (Hurst 1990)
| 1. Nincs párosodási típus, biparentális citoplazma-örökítés, a citoplazmatikus paraziták vertikálisan terjedhetnek, fittness- csökkentõk. 2. Megjelennek saját-citoplazma átadást gátló mutánsok 3. Mutáns-nem mutáns párzás
optimális (mutáns-mutáns párzás
halálos, mivel utódba nem kerül citoplazma,
nemmutáns-nemmutáns párzás alacsonyabb
fitnessû az esetleg belekerülõ paraziták
miatt) 5. Bináris rendszer |
Tehát látjuk, hogy a két nem általános, és meg is lehet indokolni. Mégis van néhány szaporodási rendszer, amelyben ennél több vagy kevesebb "nem" található. Melyek ezek az élõlények, és mik az okok?
Inkompatibilitási
rendszerek
Különbséget kell tenni a nemek (akkor nevezünk egy
párosodási típust nemnek, ha ezek fúziót
használnak) és az inkompatibilitásos párosodás-típusok
között. Ezek olyan szaporodási rendszerek, melyekben
kettõnél több típus lehet, melyek mindegyike
a többiek közül meghatározott típusokkal
párosodhat, másokkal viszont nem. Ilyen rendszert találtak
a legtöbb csillósnál, a bazídiumos gombáknál
és a nyálkagombák gametikus fúziójánál.
Ilyen párosodáskor azonban nem játszódik le
citoplazmafúzió, csak magcsere (konjugáció).
| A hypotrich csillósok pl. konjugációkor
sok párosodási típussal bírnak, míg
fúziókor csak kettõvel. Ilyenek még
az angiosperms, tunicata A peritrich csillósok elvesztették konjugációs rendszerüket, és a fúziós géncsrére tértek át, csak kétféle nem van. |
{kind=link}
{kind=link}
{kind=link}
Kettõnél kevesebb "nem" is
lehet, a Physarum multicephalum nyálkagomba plazmódiumos
(nem gametikus) fúziójánál egyáltalán
nem ismertek szaporodási típusok. Persze emellett gametikus
szaporodás is van,ahol vannak típusok.
Olyan estek is ismertek, amikor kettõnél több nem van
fúziókor is.
| Physarum multicephalum nyálkagomba
haploid gamétáinál bármelyik tud
bármelyikkel
fuzionálni, ameddig három polimorf mitokondriális
lókuszon
genetikailag nem identikusak. Mégis, a mitokondriumok
örökítõdése általában
uniparentális, Ennek a mikéntjét a polimorf
lókuszok legalább 13 allélja határozza meg,
melyek között egy dominancia-sorrend dönti el, melyik
típus mt-ja marad meg. (Mindkét gaméta viszi a
zigótába, csak az egyik féle marad meg
egyoldalú emésztési reakció során).
Tehát posztfuzionális uniparentalitás van
és mitokondriális gén
alapú inkompatibilitási rendszer. Ez esetben több nem van, de az uniparentalitás így is érvényesül. Valószinûleg azért nem találtak eddig több ilyen rendszert, mert ha az uniparentalitás a zigótaképzés után dõl el, akkor az ilyen rendszer sokkal sebezhetõbb az önzõ genetikai elemek által, mint a fúzió elõtt már eleve aszimmetrikus rendszerek (a Physarumnál tényleg van egy ilyen önzõ mt plazmid, lásd az elõzõ fejezetet). Ha az ilyen önzõ folyamatok költségesek a zigótának, akkor az a gaméta, amelyik el tudja ezt a problémát kerülni saját prefuzionális aszimmetria kifejlesztésével, az elõnyben lehet. Az esetleges több szörös nemi rendszerek így könnyen bináris típusúvá alakulhatnak. A Physarum-szerû "relatív szexualitás" ezért lehet ritka. |
Ezen kívül azonban még nem találtak olyan rendszert, ahol cp fúzió estén is kettõnél több típus lenne. Ez is a konfliktus-leszerelés fontosságát hangsúlyozza a bináris rendszerek kialakulásában.

A gaméták méretbeli differenciálódása
Az konfliktus-leszereléssel járó eltérõ párosodási típus elõnyeibõl még nem következik automatimusan a gaméták eltérõ mérete. Mi vezethet akkor az ivarsejtek annyira elterjedt és sokszor igen tekintélyes méretkülönbségéhez?
1. Parker-Baker-Smith modell:
Egyféle méretû, + és - párosodási
típusú gamétákból álló
populációban egy kis gamétákat termelõ
mutáns egyed elõnyös és elterjedhet, mert kis
befektetéssel (kicsi méret) sok proto-nõivarsejtet
tud megtermékenyíteni. (parazitálja a másik
féle sejtek befektetését). Ha viszont a zigóta
túlélési esélye nõ a gaméta
méretével, úgy egy csupa kis (+)
gamétából álló populációban
elõnyösek és elterjedhetnek a nagyobb, szesszilis gaméták.
Kétféle szelekciós hatás (diszruptív
szelekció) van, egyik nagy méretnek kedvez (jó zigótatúlélés,
fejlesztési eszköz), másik a nagy számnak (nagyobb
gamétaszám, jobb megtermékenyítési
esély, fertilizációs eszköz). Ez a szelekció
viszont csak akkor mûködhet, ha a kisebb gamétaszám
miatti hátrányt a nagyobb zigótatúlélés
elõnye túlszárnyalja.
2. A pártalálás elõsegítése
Az elõzõ modell spermakompetíciót (több
+ típus, mint -)
feltételez. Levitan szerint viszont spermalimitációs
viszonyok vezethettek az anizogámia létrejöttéhez.
Ilyen körülmények között a találkozás-segítõ
folyamatok vezethetek dimorfizmushoz.
Ilyen adaptáció lehet a petesejt növekedése,
a "nagy céltábla-biztosabb találat" elven.
|
Tengeri
uborkáknál a nagyobb tojások fittebbek, de
nagyobb találkozási ráta nem ellensúlyozza
azt, hogy kevesebb (de nagyobb)
petesejt van. |
A pártalálás módjának másféle elõsegítése lehet a spermavonzó feromonok alkalmazása. Ez esetben, ha a feromon-hatósugár arányos a termelõ sejt térfogatával, akkor megéri nagynak lenni (és nagy illatfelhõt csinálni).
3. Miért kicsi a spermium?
A PBS szerint a nagy szám miatt elõnyös kicsinek lenni.
Spermakompetícióban esetleg elõnyös lehet a
gyorsabb mozgás miatt is (de spermalimitációnál
a hosszabb életû jár jobban), másrészt
szelekció hathat a kis méretre amiatt is, hogy az organellumok
és citoplazmatikus paraziták örökíthetõsége
kicsi legyen (kevés cp legyen, lásd fejlebb).
A szex
és a fajok természete
|
Európai Alchemilla palástfüvek szűznemzők.
Kb. 300 morfológiai változata van, amik néhány klón nem
nagyon különböző változatai . (A Bdelloidea
kerekesférgek
megint renitens kivételek, mert ugyanolyan jó
morfológiai különbségeket
mutatnak, mint ivaros rokonaik). |
Ahol az ivaros fajok nem mutatnak jó elkülönülést, ott valószínűleg még telt el elég idő a földrajzi elkülönülés után.